Graptolites Radiolaria Stratigraphy
Graptolites
Graptolite Origins
The origin and evolution of coloniality in the hemichordates is a poorly understood process as fossil data are basically non-existent and the comparison of fossil Graptolithina with modern Pterobranchia is still difficult, especially as insufficient information exists on the skeletal development of living pterobranchs.
In the Graptolithina the proximal end development has been considered the most important indication for evolutionary and phylogenetic relationships. The taxonomy of the graptolites is, thus, based largely on the changes in the proximal development and the associated growth direction of the proximal (primordial) thecae. A comparison of the initial skeletal growth of modern pterobranchs and graptolites can show relationships and differences.
The initial parts of a graptolite colony are the prosicula and metasicula, while in modern – benthic – pterobranchs, the initial structure is termed a dome. The dome is formed by the secretion of the pterobranch larva and is structureless. The graptolite prosicula, however, bears distinct structures like the spiral thread (Schraubenlinie of Kraft, 1926). It is differently constructed and does not resemble the dome of a modern pterobranch.
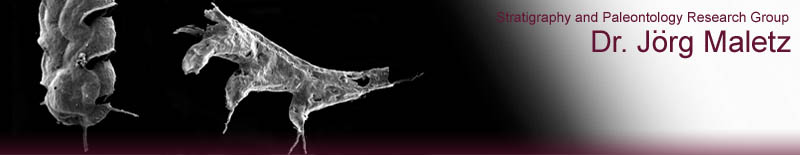
Figure
1. Initial growth of the skeletons of Rhabdopleurida (modern Pterobranchia)
and Graptolithina showing differences in development and structure (Maletz,
Steiner & Fatka 2005).
Pterobranchs are known from the Middle Cambrian to the recent, but have rarely been described from the fossil record. They are, however, common in the fossil record of certain regions, for example in the Middle Ordovician Limestones of Öland, Sweden and have been described from glacial erratic boulders in northern Germany and Poland. Numerous specimens have most likely been included in the diverse dendroid orders of the Graptolithina, while shale material may also have been identified as algae.
Dichograptid Evolution
The identification of proximal development types is essential for species identification and biostratigraphic interpretation of graptolite occurrences. Misidentification may cause considerable miscorrelations, as is known from the example of the pendent didymograptids in the Early to Middle Ordovician. Even though proximal developments tend to be conservative in dichograptids, characteristic changes can be observed and used for understanding the higher levels of taxonomy. Cladistic analysis of character associations supports the taxonomy and evolutionary studies.
The taxonomy and evolution of the Dichograptacea from the Early to Middle Ordovician has been one of my main interests for many years. The Early to Middle Ordovician faunas of Scandinavia (Sweden, Norway) and eastern North America (Quebec, western Newfoundland) are quite diverse and sometimes excellently preserved. Thus, a detailed phylogenetic analysis is possible and ongoing cladistic interpretations have started to yield promising results Carlucci & Maletz 2007, GSA Abstract).
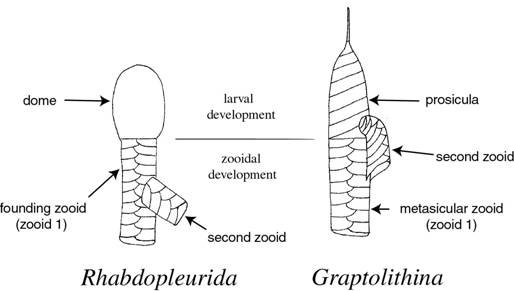
Figure
2. The proximal end development in pendent didymograptids from the Arenig
in reverse view. A. Xiphograptus mantuanus (based on Riva). B.
Didymograptellus bifidus. C. Didymograptus spinulosus. The
prosicula is shown in black (Maletz, unpublished).

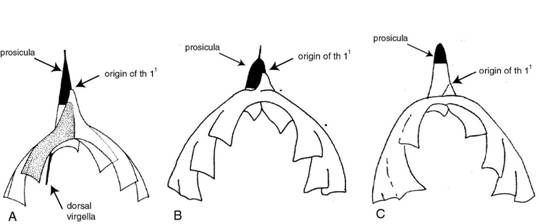
Figure 3. Relief specimens of dichograptid graptolites often show their proximal development in the reverse view. The left specimen is a latex cast of Expansograptus holmi from the Tøyen Shale at Mt. Hunneberg, Sweden. The right specimen is Didymograptus murchisoni from the Elnes Formation of the Oslo Region, Norway. The proximal development in the left specimen is of the isograptid type with a prosicular origin of the first theca, while the right specimen shows an artus type development with a low metasicular origin of the first theca. (From Toro & Maletz, 2007; GSA Poster).
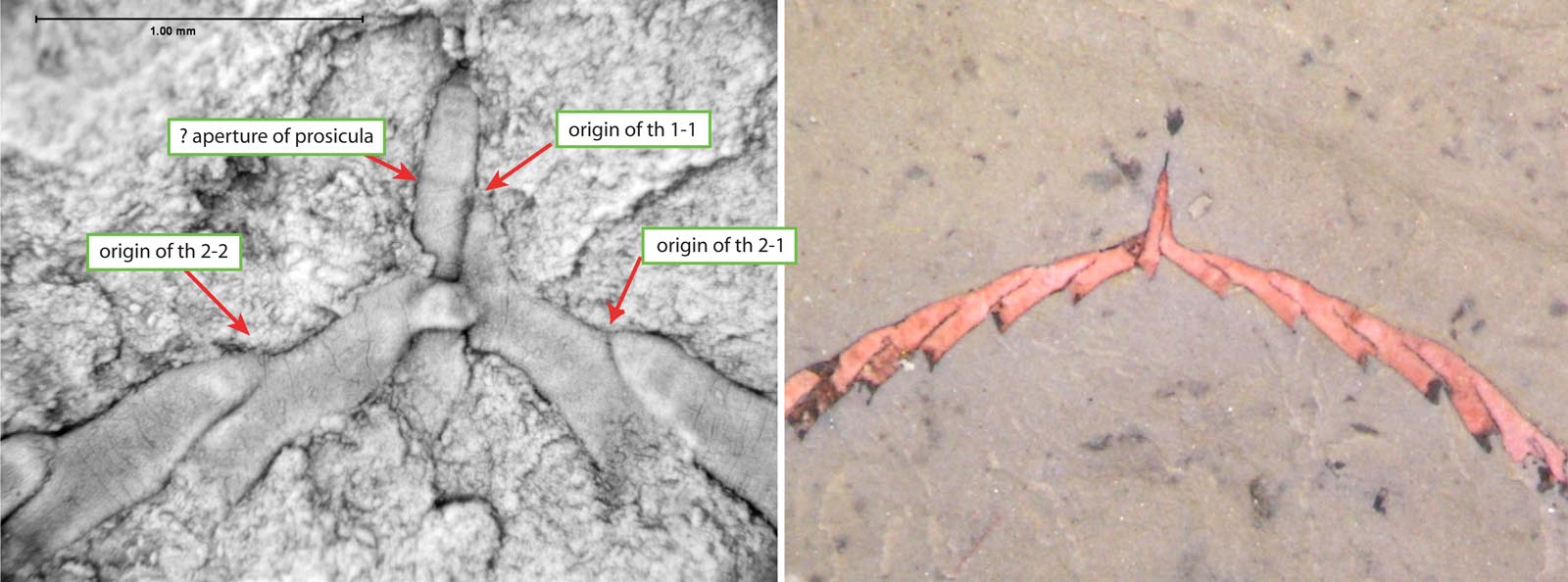
Figure 4. Cymatograptus sp. nov. This species comes from the mid-Arenig "Baltograptus deflexus" Biozone of NW Argentina. The specimens show the artus type proximal development in reverse (left) and obverse (right) views. The material is preserved in partial relief in grey mudstones of the Acoite Formation. Some of the specimens are filled with a whitish to pinkish mineral, possibly an iron oxide. Right- and left-handed specimens occur in equal numbers. Even though the material in general does not show fusellar structure, some details can be inferred. The specimen on the left shows the dimensions of the prosicula through a slight constriction at the boundary to the metasicula. The specimens were provided by Blanca Toro (Mendoza, Argentina).
Graptolite Cladistics
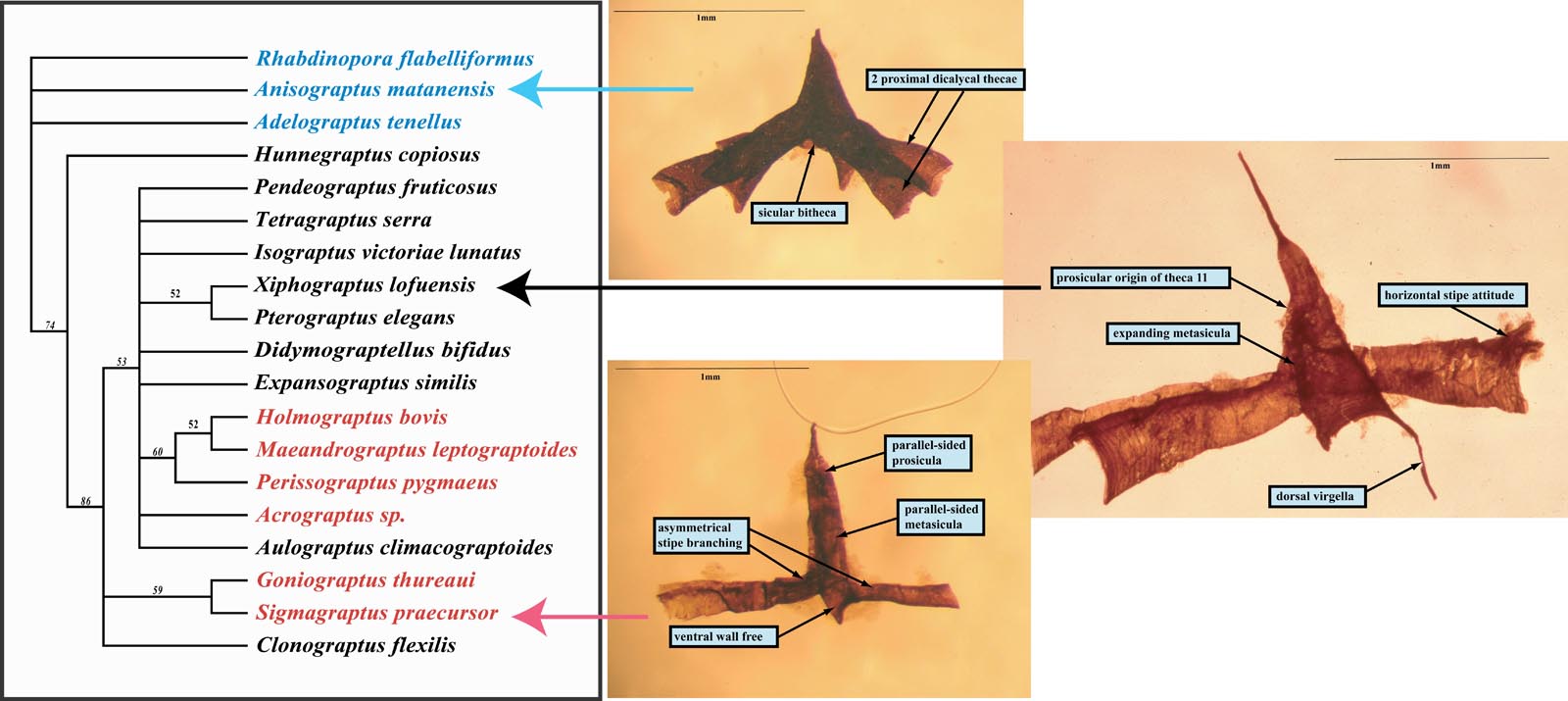
Figure 5. Preliminary cladistic interpretation of the main Dichograptacean groups. Outgroup Anisograptidae (blue); Dichograptidae (black); Sinograptidae (red). Based on Carlucci & Maletz (2007, GSA Poster).
Paleogeography
In many cases, the paleogeographic distribution of graptolites is useful for biogeographic interpretations and plate tectonic reconstructions. In general, Graptolites show a differentiation from cold water faunas (Atlantic Faunal Province) to warm water faunas (Pacific Faunal Province) in the Ordovician. Also, shallow water and deep water faunal elements can be differentiated and used to outline continental shelves. Research in Scandinavia (Atlantic Faunal Province) and eastern North America (Pacific Faunal Province) helps to understand the faunal differentiation as well as individual ecological requirements of faunal elements. The investigation also includes studies on the faunal diversities and changes through time.
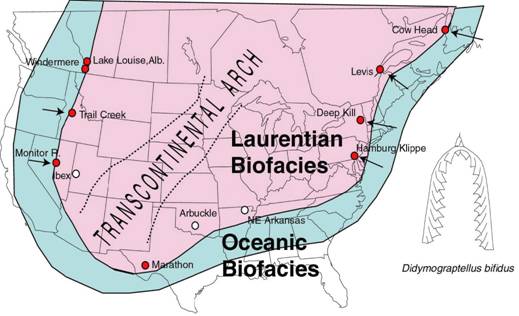
Figure
6. The paleogeographic distribution of the Pacific Faunal Province element
Didymograptellus bifidus in the mid Arenig of North America (Laurentia).
Arrows show where lithological successions are thrusted onto the
Laurentian platform during later orogenies. The Laurentian biofacies
refers to the shallow water shelf faunas, while the oceanic biofacies
indicates deep water graptolite faunas (Maletz, unpublished).
Biostratigraphy
Graptolites are the most important faunal elements for biostratigraphic purposes in the early Palaeozoic. They are used to define Global stratotype sections (GSSP’s) for a number of chronostratigraphic time intervals from the early Ordovician to the Devonian.
The isograptids in the mid-Arenig to mid-Darriwilian especially, show promise for both a phylogenetic analysis and a high potential for biostratigraphic purposes as their proximal structures and rhabdosome shapes changes quickly through time. Still, they are easily identified and differentiated when well preserved.
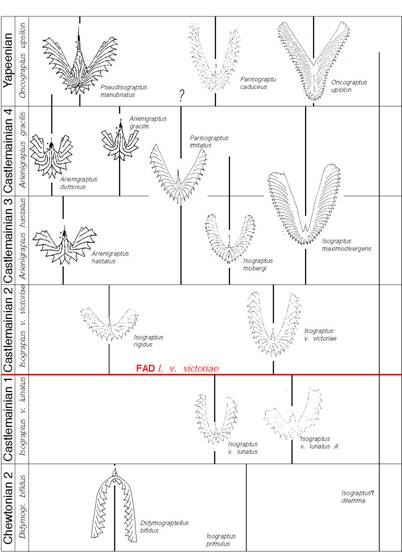
Figure
7. Isograptid biostratigraphy in the Chewtonian to Yapeenian (early Middle
Ordovician) of the Cow Head Group of western Newfoundland, one of the
standard – and most complete - successions for Middle Ordovician
isograptids and pseudisograptids (Maletz, unpublished).
Evolution of the Diplograptina
The early Middle Ordovician isograptids evolved into the most important Ordovician faunal elements: the Diplograptaceans, during the late Arenig time interval. These biserial graptolites show extremely complex proximal developments that can be figured out only through investigation of relief specimens and series of isolated growth stages, a work that is mainly done by SEM photography today. However, transmitted light investigation can still provide important data used to help untangle the complex interactions of the thecal tubes with the use of their fusellar structure.
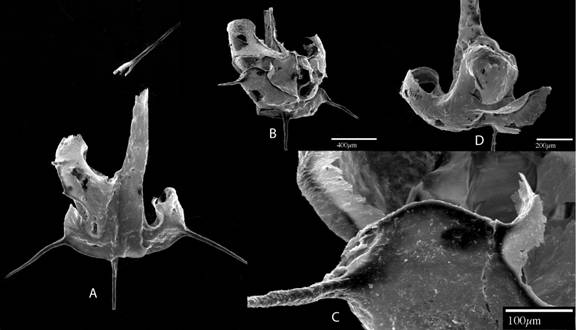
Figure
8. Juvenile specimens of biserial graptolites show important characters to
interpret their proximal development. A: Dicranograptus sp.,
Viola, Limestone. B, C, D. Archiclimacograptus sp., Table Head Group,
western Newfoundland. SEM photos (Maletz, unpublished).
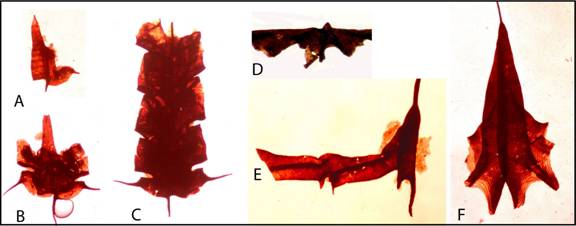
Figure
9. Isolated and bleached graptolites from shales and limestones showing
fusellar structure. A-C. Archiclimacograptus sp., Table Head Group,
flattened specimens, growth series. D. Expansograptus abditus,
relief, Cow Head Group. E. Nicholsonograptus fasciculatus,
flattened, no fusellar structure visible, Table Head Group. F.
Isograptus victoriae lunatus, flattened, Cow Head Group. All specimens
from western Newfoundland. Transmitted light photographs (Maletz,
unpublished).
Silurian Monograptids
It is often difficult to identify Silurian monograptids, as their elaborate thecal apertures are flattened on shale surfaces and details are not visible any more. Thus, isolated specimens are highly sought after in order to help understand the structure, but are known from few regions. In the Llandovery of Dalarna, central Sweden, limestone lenses can be collected from the Kallholn Shale, which bares a diverse fauna of Llandovery biserial and uniserial (monograptid) graptolites.
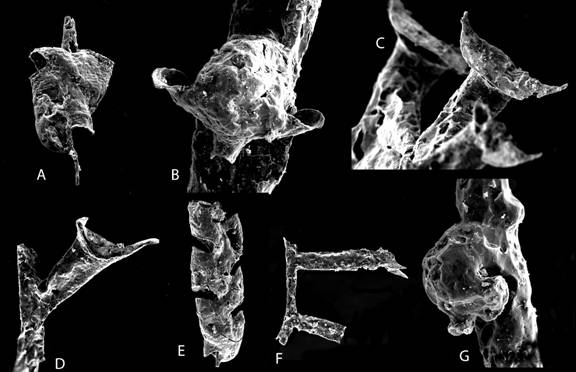
Figure
10. Llandovery graptolites from Dalarna, Sweden. A. Normalograptus
scalaris, juvenile in obverse view. B, G. thecal aperture of
Streptograptus dalecarlicus, proximal (G) and distal (B) thecal
apertures. C. Oktavites contortus, thecal apertures. D, F.
Lituigraptus convolutus, thecal aperture and rhabdosome fragment. E.
Metaclimacograptus sp., reverse view (Maletz, unpublished).
Retiolitid rhabdosome architecture
In the Silurian, one group of biserial graptolites, the retiolitids, looses the typical periderm seen in most graptolites. These species are preserved only as a meshwork of lists that can be shown to be formed originally on a membrane surface. Unfortunately, this membrane is rarely preserved in the fossil record. The reason for the list development is unknown. It can make identification of material extremely difficult and, as with many specimens, it is severely distorted by post-mortem preservational effects. The taxonomy and biostratigraphy of the retiolitids remains difficult, but exciting work.
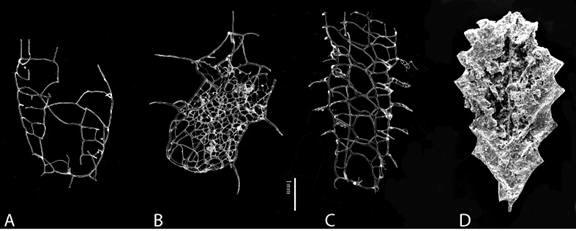
Figure
11. SEM photos of Silurian biserial graptolites. A. Plectograptus
macilentus. B. Spinograptus spinosus. C. Plectograptus
robustus.
D. Petalolithus sp. A-C are retiolitids from glacial erratic
boulders, collected in northern Germany, specimens isolated by Hermann
Jaeger, Berlin, Museum für Naturkunde. D is an isolated specimen from the
Kallholn Shale in the Llandovery of Dalarna, Sweden (Maletz, unpublished).
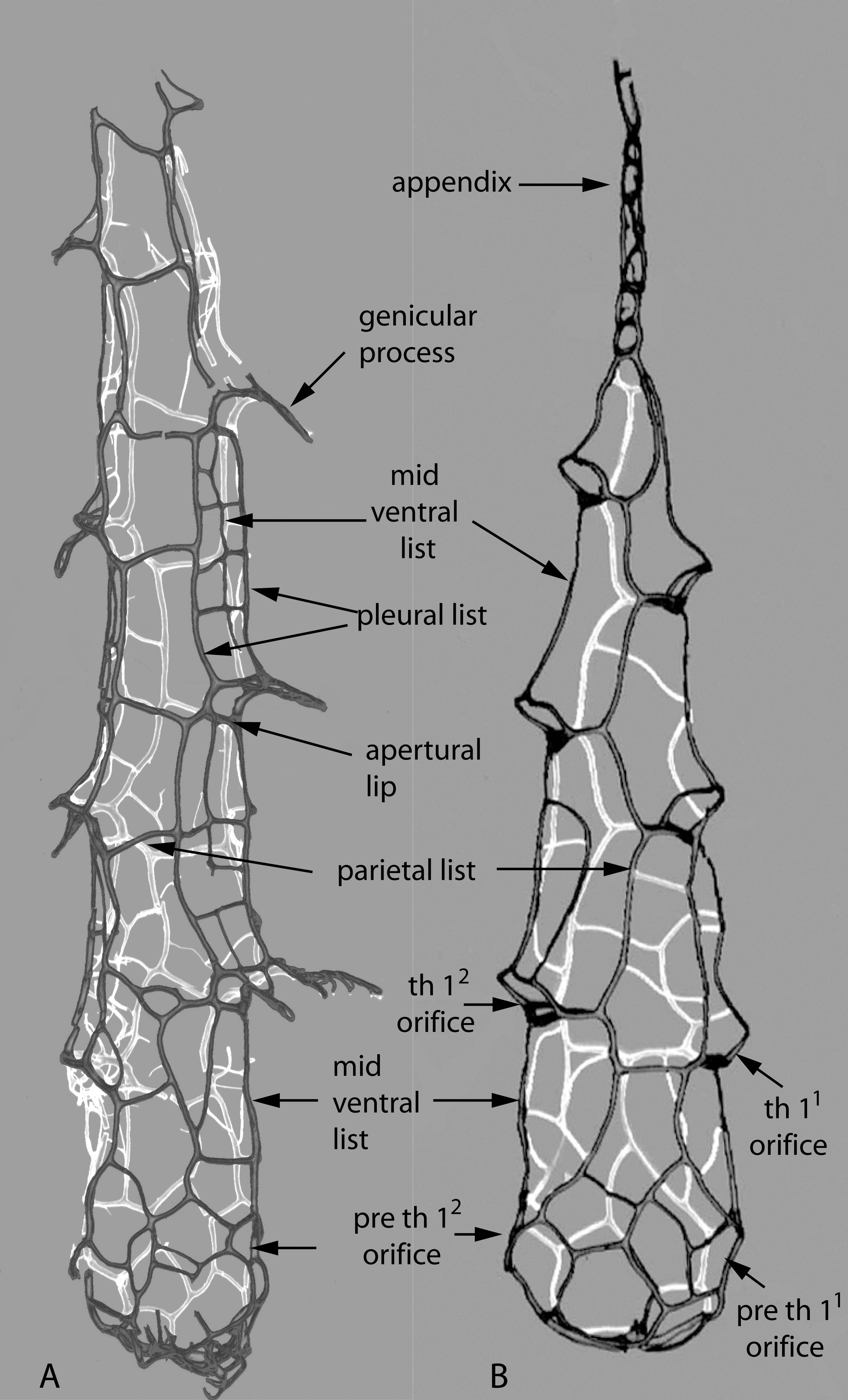
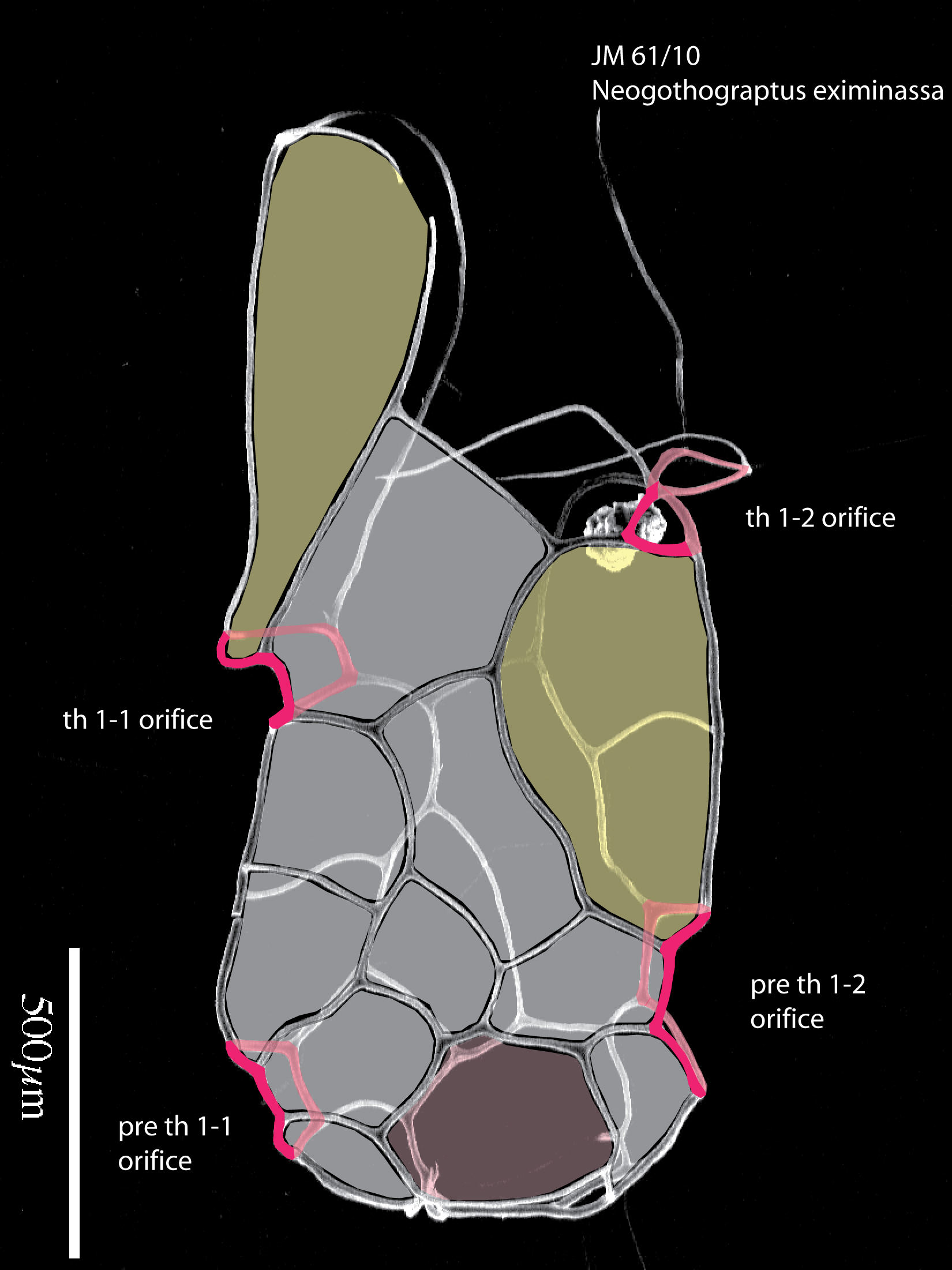
Figure 12. A. Neogothograptus eximinassa. B. Holoretiolites erraticus. Illustrations showing the list structure of these two closely related genera, frontal lists dark, back lists shown in white. C. Neogothograptus eximinassa, juvenile specimen with indications of ancora sleeve (in grey and yellow); ancora sleeve orifices indicated in red. Material from the Jaeger collection (Museum für Naturkunde, Berlin).